Nervenzelle, Reizweiterleitung, Reizverarbeitung
Das Nervensystem besteht wie alle anderen Organe aus einzelnen Zellen. Diese sind aber gänzlich anders aufgebaut als andere Körperzellen. Sie sind geradezu perfekt an ihre Aufgabe der Informationsleitung und -verarbeitung angepasst – sie sind hochspezialisiert. Diese Spezialisierung führt aber auch dazu, dass sie nicht mehr in der Lage sind sich zu teilen, d.h. sich zu vermehren oder abgestorbene Zellen zu ersetzen. „Wunden“ im Nervensystem können nicht heilen!
Nervenzellen können sehr unterschiedliche Formen und Größen haben, dennoch besitzen sie alle einen gemeinsamen Grundbauplan. Sie bestehen aus einem Zellkörper, dem Soma, das zwischen 0,01 und 0,1 mm groß sein kann. Hier liegen der Zellkern und eine große Anzahl an Mitochondrien, kleinen Energiekraftwerken in der Zelle.
Vom Zellkörper zweigen zahlreiche Verbindungen ab, die den Kontakt zu anderen Nervenzellen herstellen, um Informationen aufzunehmen und weiterzuleiten. Man weiß heute, dass eine einzige Nervenzelle mit bis zu 10.000 anderen Nervenzellen in Verbindung stehen kann. Dabei läuft der Informationsfluss immer nur in eine Richtung. Ankommende Reize werden von den Dendriten aufgenommen, kabelartigen Fortsätzen, die man sich wie Antennen vorstellen kann. Diese leiten die Reize zum Zellkörper. Hier treffen die Reize aus den einzelnen Dendriten zusammen und erregende und hemmende Eingänge werden miteinander verrechnet (Summation). Überwiegt die Erregung, so wird auch diese Zelle „feuern“, d.h. die Information an andere Nervenzellen, Muskeln oder Drüsen weitergeben. Überwiegt aber die Hemmung, so wird sie schweigen.
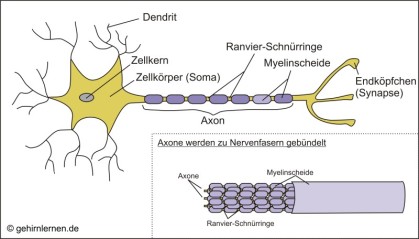 Abb. 10.1: Aufbau der Nervenzelle
Abb. 10.1: Aufbau der Nervenzelle
Die Erregungsweiterleitung geschieht über das Axon. Axone können bei einem Durchmesser von nur 0,002 bis 0,01mm bis zu 1m lang sein!
An jedem Dendriten und jedem Axon findet sich am Ende ein kleines Köpfchen, die Synapse. Über diese wird die Verbindung zu anderen Nervenzellen oder Körperzellen hergestellt. Dazu aber später mehr.
Wie funktioniert diese Reizweiterleitung?
Die Reizweiterleitung erfolgt in Form von elektrischen Impulsen. Wie aber kommt Strom in eine Nervenzelle? Durch das Membranpotential, das jede Körperzelle besitzt. In der Nervenzelle nennt man es Ruhepotential (im Gegensatz zum Aktionspotential bei der Reizweiterleitung).
Dieses wird durch Ladungsträger, Ionen, aufgebaut, die sich in bestimmten Konzentrationen innerhalb und außerhalb der Zelle befinden (Abb. 10.2). Dabei findet sich in der Zelle ein Überschuss an negativen, außerhalb der Zelle ein Überschuss an positiv geladenen Ionen.
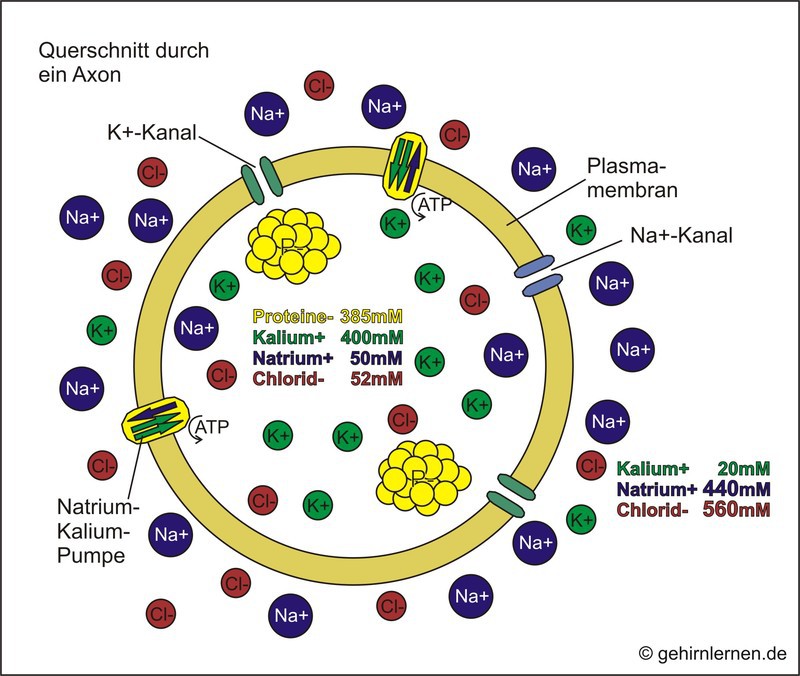 Abb. 10.2: Ionenverteilung beim Ruhepotential
Abb. 10.2: Ionenverteilung beim Ruhepotential
Der schematische Querschnitt durch ein Axon zeigt die Ionenverteilung innerhalb und außerhalb der Zelle. Innen findet sich ein Überschuss an negativ geladenen Proteinen und positiv geladenen Kalium-Ionen (K+). Außen findet sich hingegen ein Überschuss an negativ geladenen Chlorid-Ionen (Cl-) und positiv geladenen Natrium-Ionen (Na+).
Wie wird dieses Ungleichgewicht aufgebaut und welche Bedeutung hat es?
Ionen sind in wässriger Lösung von einer Wasserhülle umgeben. Da die Membran aus Fettmolekülen aufgebaut ist, können die Ionen nicht einfach hindurchdiffundieren, sie müssen transportiert werden, um das oben genannte Ungleichgewicht von positiven und negativen Ladungen herzustellen. Dafür hat die Zelle zwei Mechanismen:
Tunnelproteine in der Membran, die so genannten Ionenkanäle, erlauben einen Durchtritt bestimmter Ionen durch die Membran und ermöglichen so die Aufrechterhaltung oder Änderung der Spannung an der Membran. Es gibt verschiedene Arten von Kanälen:
-
Kanäle, die immer offen sind
-
Spannungsgesteuerte Kanäle, die sich abhängig von den Außenbedingungen öffnen und schließen
-
Ligandengesteuerte Kanäle, die sich nur öffnen, wenn ein bestimmtes Molekül (z.B. ein Neurotransmitter) an sie bindet
-
Mechanisch gesteuerte Kanäle, die sich – wie z.B. die Tastsinneszellen der Haut – auf mechanische Belastung hin öffnen oder schließen
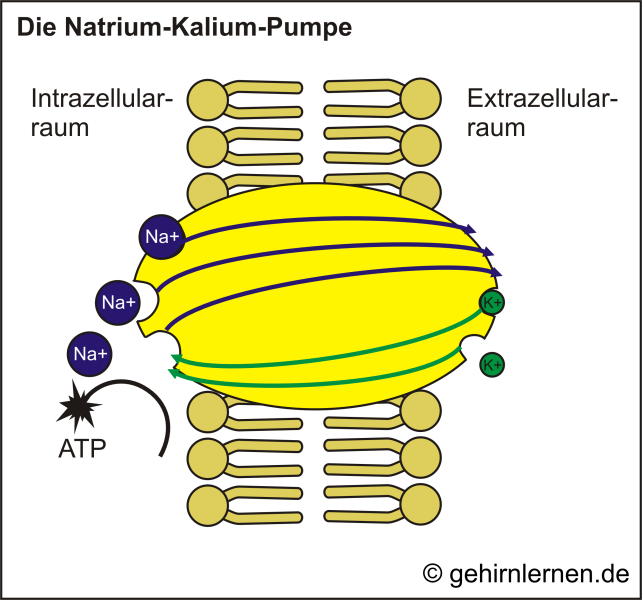 Abb. 10.3: Die Natrium-Kalium-Pumpe
Abb. 10.3: Die Natrium-Kalium-Pumpe
Kanäle können nur das passive Diffundieren von Ionen zu-lassen, nicht aber aktiv Ionen gegen das Konzentrations-gefälle pumpen. Um das Ungleichgewicht von Kalium-Ionen und Natrium-Ionen herzustellen, bedarf es somit eines aktiven Transportes. Dieser spezielle Transport-mechanismus heißt Natrium-Kalium-Pumpe (Abb. 10.3). Unter Energieverbrauch (d.h. (für Profis) ATP-Spaltung) pumpt diese positiv geladene Kaliumionen (K+) in die Zelle hinein und positiv geladene Natriumionen (Na+) aus der Zelle heraus. Dadurch wird in der Zelle ein K+-Überschuss, außerhalb der Zelle ein Na+-Überschuss erzeugt. Da aber einige K+-Kanäle in der Membran immer geöffnet sind, können K+-Ionen ihrem Konzentrationsgefälle folgend aus der Zelle hinaus diffundieren. Da im Gegenzug aber nur sehr wenige Na+-Kanäle offen sind, können kaum Na+-Ionen zum Ausgleich hineinströmen. Das Zellinnere verliert somit positive Ladungen und es entsteht eine negative Spannung von ca. -60mV (Abb. 10.4).
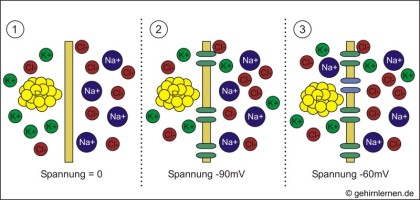 Abb. 10.4: Entstehung des Ruhepotentials
Abb. 10.4: Entstehung des Ruhepotentials
Die Natrium-Kalium-Pumpe schafft ein Ungleichgewicht mit einem Überschuss an K+-Ionen im Innern und einem Na+-Überschuss im Äußeren der Zelle (1). Offene K+-Kanäle in der Membran ermöglichen die Diffusion von K+ seinem Konzentrationsgefälle folgend aus der Zelle hinaus. Gäbe es (rein hypothetisch) nur K+-Kanäle so würde eine Spannung von -90mV entstehen (2). Da es aber auch einige wenige Na+-Kanäle gibt, können zum Ausgleich auch einige Na+-Ionen wieder in die Zelle hineindiffundieren. So pendelt sich die Spannung bei ‑60mV, dem Ruhepotential der Nervenzelle, ein (3).
Das Aktionspotential
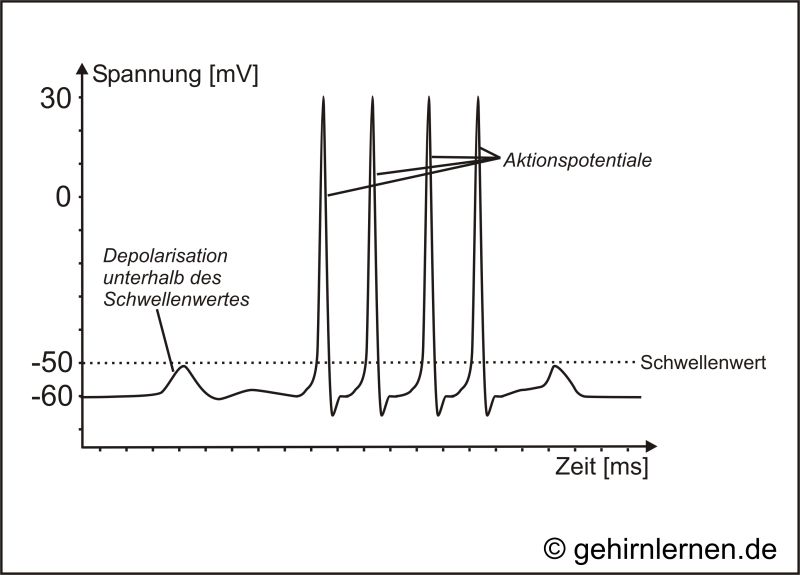 Abb. 10.5: Schema der Ableitung eines Aktionspotentials an einem Axon
Abb. 10.5: Schema der Ableitung eines Aktionspotentials an einem Axon
Reize können das Membranpotential einer Nervenzelle verändern. Wird eine Nervenzelle leicht gereizt, so wird das Membranpotential etwas weniger negativ als das Ruhepotential. Man bezeichnet dies als Depolarisation. Überschreitet die Depolarisation einen bestimmten Schwellenwert von -50mV, so wird die Membran sehr rasch und kurzfristig auf +30mV depolarisiert (Abb. 10.5). Diese Reaktion wird als Aktionspotential bezeichnet. Dieses dauert nur etwa 1 bis 2 ms, dann folgt eine schnelle Rückkehr zum Ruhepotential, wobei die Membran kurzzeitig hyperpolarisiert werden kann, d.h. dass das Membranpotential negativere Werte als das Ruhepotential annehmen kann (Abb. 10.6).
Verantwortlich für das Aktionspotential sind spannungsgesteuerte Na+- und
K+-Kanäle in der Membran des Axons (Da diese nur im Axon zu finden sind, kann auch nur im Axon, nicht aber in den Dendriten, ein Aktionspotential ausgelöst werden.).
Beim Ruhepotential sind alle spannungs-gesteuerten Kanäle geschlossen. Wird das Axon über den Schwellenwert von -50mV depolarisiert, so öffnen sich die spannungsgesteuerten Na+-Kanäle in
der Membran und erlauben einen Einstrom von Na+ in die Zelle, wodurch das Zellinnere kurzfristig positiver wird als der Außenraum (+30mV). Bereits nach 1 bis 2ms schließen sich die Na+-Kanäle wieder,
auch wenn die Depolarisation noch andauert. Danach bleiben die Na+-Kanäle ungeachtet möglicher weiterer Depolarisationen für 1 bis 2 ms geschlossen. In dieser Phase kann kein neues
Aktionspotential ausgelöst werden. Diese Zeit nennt man Refraktärzeit. Auch spannungsgesteuerte K+-Kanäle werden durch die Depolarisation geöffnet. Diese Kanäle sind aber so
„träge“, d.h. sie öffnen und schließen sich so langsam, dass sie ihren offenen Zustand erst erreichen, wenn die Na+-Kanäle sich bereits wieder zu schließen beginnen. K+ kann nun aus der Zelle
ausströmen und so das Membranpotential wieder senken. Aufgrund der Trägheit der K+-Kanäle strömt kurzfristig sogar mehr K+ aus, als für die Wiederherstellung des Ruhepotentials nötig wäre, so
dass das Membranpotential kurzfristig unter das Ruhepotential absinkt.
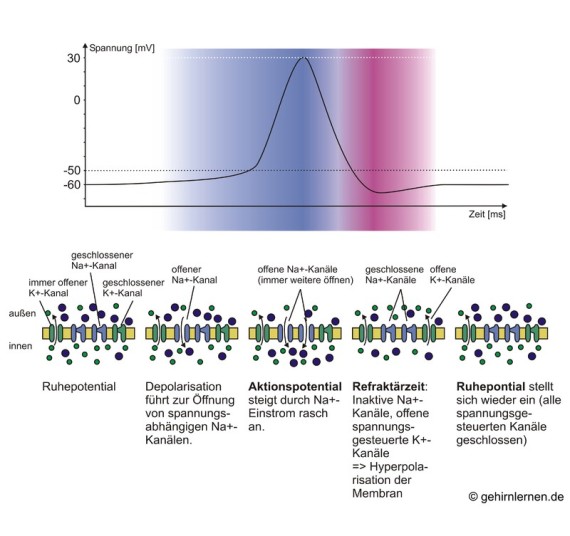 Abb. 10.6: Ablauf des Aktionspotentials
Abb. 10.6: Ablauf des Aktionspotentials
Ein Aktionspotential macht noch keine Reizweiterleitung
Wenn aber an einer Stelle ein Aktionspotential entsteht, bedeutet dies, dass an dieser Stelle das Innenmedium plötzlich positiv geladen ist. In der Nachbarschaft herrschen aber weiterhin negative Ladungen vor. Da gegensätzliche Ladungen sich bekanntlich anziehen, verschieben sich die Ionen und depolarisieren die Nachbarbereiche am Axon mit. Erreicht diese Depolarisation den Schwellenwert, wird auch hier ein Aktionspotential ausgelöst.
Warum läuft das Aktionspotential dann nicht in beide Richtungen vom Ursprung des ersten Aktionspotentials weg? Das Aktionspotential kann nur in eine Richtung verlaufen, da die „hinter“ dem Aktionspotential liegenden Bereiche noch in der Refraktärzeit (s.o.) liegen, hier also noch kein Aktions-potential wieder ausgelöst werden kann.
Diese Erregungsweiterleitung verläuft relativ langsam, da die Verschiebung der Ionen innerhalb des Axons durch das Vorhandensein der Ladungen im Außenmedium gebremst wird. Darum gilt auch: je dicker ein Axon, desto schneller die Reizweiterleitung – genau wie bei einem Stromkabel: je dicker, desto geringer der Widerstand.
Wirbeltiere haben zudem an etwa der Hälfte ihrer Nervenzellen eine Isolierung um die Axone, um die Reizweiterleitung zu beschleunigen. Diese Isolierung wird von Gliazellen gebildet, den Stützzellen des Nervensystems. Sie besteht aus Myelin und wird darum auch Myelinscheide genannt (vgl. Abb. 10.1). Aktionspotentiale können an diesen Axonen ausschließlich in Einschnürungen der Myelinscheide, den Ranvier-Schnürringen, entstehen, da nur hier Na+-Kanäle zu finden sind. Die Ausbildung eines Aktions-potentials führt zur Depolarisation der Membran im nächsten Schnürring, so dass das Aktionspotential sozusagen von Schnürring zu Schnürring springt. Darum spricht man auch von saltatorischer Reizweiterleitung (saltare – lat. springen, tanzen). Dabei läuft das Aktionspotential am myelinierten deutlich schneller als am unmyelinierten Axon, da die (oben genannten) Ionen sich Dank der Isolierung und damit der abgeschirmten Außenladungen schneller verschieben.
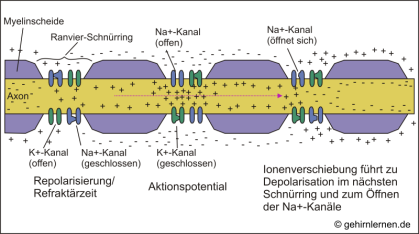 Abb. 10.7: Die saltatorische Reizweiterleitung
Abb. 10.7: Die saltatorische Reizweiterleitung
Ein Aktionspotential in einem Ranvier-Schnürring (hier in der Mitte dargestellt) führt zu Ionenverschiebungen, die die Membran in den benachbarten Schnürringen depolarisieren. "Hinter" dem momentanen Aktionspotential (links dargestellt) kann aufgrund der Refraktärzeit kein Aktionspotential ausgelöst werden. Im nächsten Schnürring (rechts dargestellt) führt die Depolarisation zur Auslösung eines weiteren Aktionspotentials. So springt das Aktions-potential von einem Ranvier-Schnürring zum nächsten entlang des Axons (saltatorische Reizweiterleitung).
Die Kommunikation zwischen den einzelnen Nervenzellen
Irgendwann ist das Ende des Axons erreicht. Um die weitere Erregungsweiterleitung zu ermöglichen, muss die Erregung nun an die nächste Nervenzelle – genauer gesagt ihre Dendriten – weitergegeben werden. Dies geschieht an den Verbindungsstellen, den Synapsen. Diese bestehen aus dem Endköpfchen des Axons der Senderzelle (Präsynapse, weil sie vor der Synapse liegt) und dem Endköpfchen des Dendriten der Empfängerzelle (Postsynapse, weil sie hinter der Synapse liegt). Den Raum zwischen Prä- und Postsynapse nennt man synaptischen Spalt. Man unterscheidet zwei Übertragungswege und damit Formen von Synapsen.
- Elektrische Synpasen
- Chemische Synapsen
Bei den elektrischen Synapsen liegen Prä- und Postsynapse so dicht bei einander und sind mit kleinen Brücken (gap junctions) verbunden, dass das Aktionspotential ungehindert von einer auf die andere Nervenzelle überlaufen kann. Diese Form der Übertragung ist sehr schnell, die Erregung kann aber nur 1:1 von einer auf die andere Zelle übertragen werden.
Chemische Synapsen kommen weitaus häufiger im Nervensystem vor. Bei ihnen wird das elektrische Signal in ein chemisches Signal umgewandelt, das so den synaptischen Spalt überquert. An der Membran der Postsynapse wird das chemische Signal dann wieder in ein elektrisches Signal umgewandelt.
Dieser Übertragungsweg ist zwar etwas langsamer, ermöglicht aber durch den Einsatz unterschiedlicher, chemischer Botenstoffe (Neurotransmitter) die Modulation des Signals. Botenstoffe können nämlich sowohl eine Erregung als auch eine Hemmung der postsynaptischen Nervenzelle hervorrufen.
In diesem Beispiel wird ein erregendes postsynaptisches Potential ausgelöst (EPSP). Neben diesen erregenden Synapsen gibt es im Nervensystem aber auch hemmende Synapsen. Der wichtigste hemmende Botenstoff ist GABA. Die GABA-Rezeptoren an der postsynaptischen Membran sind nicht mit Na+/K+-Kanälen gekoppelt sondern mit K+- und Chlorid(Cl-)-Kanälen. Bindet GABA an diese Rezeptoren, öffnen sich die Kanäle und K+-Ionen strömen aus, während Cl- in die Zelle einströmen kann. Dadurch wird das negativ geladene Zellinnere noch negativer, die Membran wird auf -80mV hyperpolarisiert. Man spricht von einem inhibitorischen postsynaptischen Potential (IPSP).
Wie wird das elektrische Signal in ein chemisches und dann wieder in ein elektrisches umgewandelt?
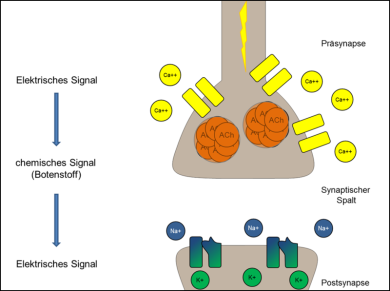
Betrachten wir zunächst ein einfaches Beispiel einer cholinergen Faser, die ein Signal auf eine Muskelfaser (also keine weitere Nervenzelle – darum „einfaches Beispiel“), die so genannte muskuläre Endplatte, überträgt. Erreicht ein Aktionspotential das Endköpfchen, also die Präsynapse, so öffnen sich hier in der spannungsgesteuerte Calcium++(Ca++)-Kanäle, durch die Ca++ in das Zellinnere einströmen kann. Dieser Einstrom bewirkt eine Verschmelzung kleiner Bläschen, in denen sich der Neurotransmitter Acetylcholin befindet, mit der präsynaptischen Membran. Diese so genannten Vesikel platzen und geben den Neurotransmitter in den synaptischen Spalt frei. Er diffundiert in kaum mehr als 0,1ms durch den Spalt und bindet in der postsynaptischen Membran an für ihn spezifische Bindungsstellen, die Rezeptoren. Diese Rezeptoren sind direkt an einen Ionenkanal gekoppelt, der sich durch das Andocken von Acetylcholin an den Rezeptor öffnet (ligandengesteuerter Kanal (s.o.)). Dieser Ionenkanal ist im Falle von Acetylcholin sowohl für Na+-Ionen als auch geringfügig für K+-Ionen durchlässig. Als Folge der Aktivierung durch Acetylcholin strömt also viel Na+ in die Zelle ein und nur wenig K+ aus. Wie bei einer Nervenzelle wird die Muskelzelle dadurch depolarisiert. Für die Muskelzelle bedeutet diese eine Aktivierung und somit eine Kontraktion der Muskelfaser.
Die Abläufe an der chemischen Synapse können Sie sich über folgenden Link auch als Video ansehen:
Informationsverarbeitung
Jede einzelne Nervenzelle steht aber mit vielen erregenden und hemmenden Synapsen in Verbindung. Werden mehrere EPSPs gleichzeitig an verschiedenen Synapsen einer Zelle ausgelöst, so summieren sich diese und können am Axonursprung zu einer Depolarisierung über den Schwellenwert und somit zu einem Aktionspotential führen (räumliche Summation). Gleiches gilt, wenn mehrere EPSPs kurz hintereinander an einer Synapse ausgelöst werden (zeitliche Summation). Die Veränderung des Potentials am Axonursprung ist also die Summe aller zu diesem Zeitpunkt „einlaufenden“ erregenden und hemmenden Eingänge, die dann in ein auslaufendes Aktionspotential übersetzt werden.
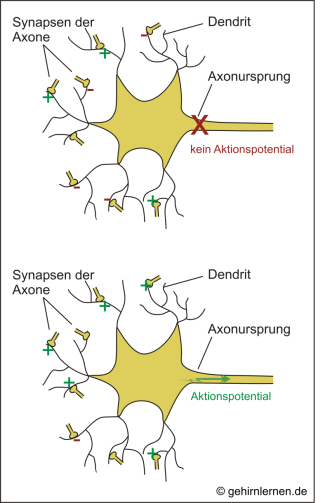 Abb. 10.8: Summation
Abb. 10.8: Summation
Erregende und hemmende Eingänge werden am Axonursprung verrechnet (Summation) und dann in ein Aktionspotential oder kein Aktionspotential übersetzt.