Das limbische System oder das „Säugergehirn"
Oberhalb des Hirnstamms schließt sich das limbische System an. In der Evolution entstand das Limbische System in der Phase der Entwicklung der Säugetiere. Darum wird es auch als Säugerhirn bezeichnet, da es allen Säugetieren gemein ist. Es reguliert die für die soziale Natur der Säugetiere typischen Empfindungen wie Sorge um den Nachwuchs, Angst, Liebe, Lust, Spieltrieb und das Lernen durch Nachahmen.
Seinen Namen erhielt das limbische System von Paul Broca («la grande lobe limbique»), der es 1878 zum ersten Mal beschrieb. Dabei bildet das Limbische System eigentlich keine anatomische, sondern eher eine funktionale Einheit basierend auf den Funktionen von Emotion, Antrieb und Lernen.
Im Zentrum des limbischen Systems steht der Papez-Kreislauf (1937), benannt nach seinem Entdecker James W. Papez. Papez meinte damit - wie auch heute fälschlicherweise noch weit verbreitet ist - einen Erregungskreislauf für Emotionen zu beschreiben. Heute weiß man aber, dass der Papez-Kreislauf vielmehr von elementarer Bedeutung für die Gedächtnisbildung ist (auch wenn in manchen Büchern (leider) immer noch etwas anderes steht…).
Seinen Ursprung (sofern man bei einem Kreislauf überhaupt von einem Ursprung sprechen kann) nimmt der Papez-Kreislauf im Hippocampus. Von dort zieht er über den Fornix zu den Mamillarkörperchen im Hypothalamus. Von dort geht es weiter zum cingulären Cortex und zurück zum Hippocampus.
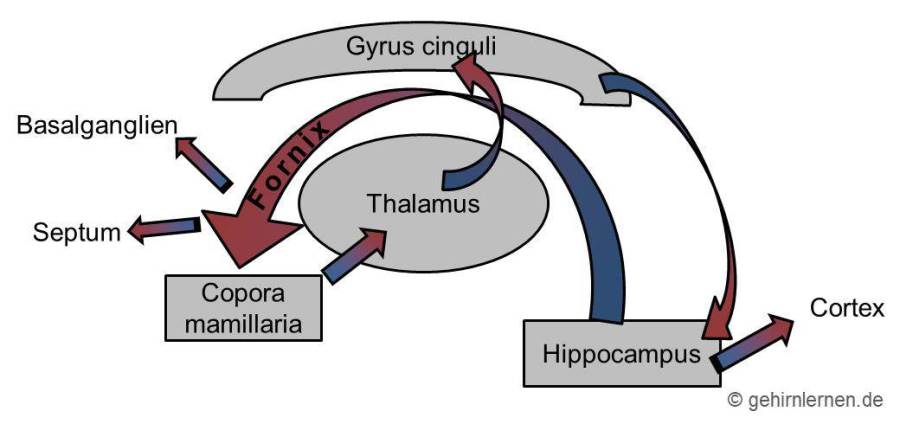 Abb. 5.1 Papez-Schleife
Abb. 5.1 Papez-Schleife
Unter funktionalen Gesichtspunkten werden heute außerdem die Amygdala, der Nucleus accumbens und Teile des Riechhirns zum Limbischen System gezählt.
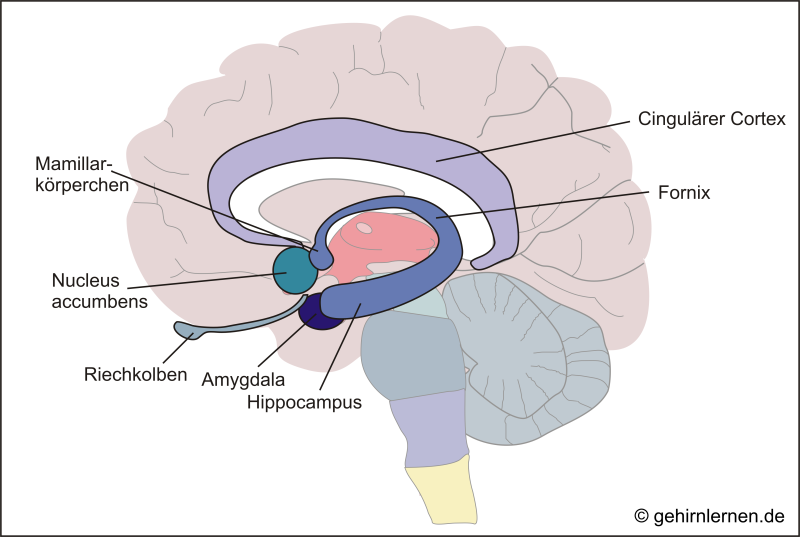 Abb. 5.2: Das limbische System
Abb. 5.2: Das limbische System
|
Inzwischen weiß man, dass noch weitere Hirnstrukturen bei der Entstehung von Emotionen, Antrieb und Gedächtnisbildung eine Rolle spielen (z.B. Kerne im Septum, Indusium griseum u.a.). Spricht jemand vom Limbischen System, so kann man eigentlich nie sicher sein, welche Gehirnareale er damit meint, wohl aber, dass er von den Funktionen „Emotion“, „Antrieb“ und „Lernen“ spricht. |
Dies verdeutlicht sehr schön das „Problem“ funktionaler Beschreibungen im Gehirn: Im Prinzip werden alle Funktionen des Gehirns von einem komplizierten Zusammenspiel aller Gehirnareale unter Rückkopplung mit dem Körper hervorgebracht, nie nur von einem einzelnen Areal allein! |
Der Hippocampus
Der Hippocampus ist eine wurmartig gebogene Struktur, die innen am Schläfenlappen (Temporallappen) unterhalb des Seitenventrikels liegt. Vorn (frontal) grenzt sie an die Amygdala im Temporallappen, hinten (caudal) an das hintere Ende des Balkens (Corpus callosum), wo sie in den Fornix, einen Faserbogen, der die meisten Ausgänge des Hippocampus führt, übergeht.
Sein Name bedeutet übersetzt „Seepferdchen“, weil irgendjemand einmal glaubte, er sähe anatomisch aus wie ein Seepferdchen. Darüber kann man sich gewiss streiten – nicht aber über seine Funktion: dem Hippocampus kommt eine entscheidende Rolle bei der Gedächtnisbildung zu. Wer keinen Hippocampus hat, der kann keine neuen Erinnerungen mehr abspeichern. Zum Glück hat aber jeder zwei – in jeder Hirnhälfte einen – und der Verlust eines Hippocampus ist durchaus zu verschmerzen.
Der Hippocampus ist eine corticale Struktur, d.h. er weist genau wie der übrige Cortex eine plattenartige Schichtung auf. Allerdings besteht diese Schichtung im Hippocampus nur aus drei statt aus sechs Schichten. Das hat damit zu tun, dass der Hippocampus ein evolutionär sehr altes Gebiet des Cortex ist. Darum spricht man auch von Allocortex (=anderer Cortex) oder Archicortex (=alter Cortex). Typisch für den Hippocampus ist zudem, dass diese drei Schichten wie bei einer Biskuitrolle in einander gerollt sind.
Heute weiß man, dass der Hippocampus keine gleichförmige Struktur ist, sondern aus mehreren Strukturen besteht. Wenn man es genau nimmt, spricht man darum lieber von der Hippocampus-Formation oder hippocampalen Formation. Die einzelnen Strukturen sind
- der Hippocampus proper (eigentlicher Hippocampus),
- der Gyrus dentatus und
- das Subiculum.
Manche zählen auch noch den entorhinalen Cortex zur hippocampalen Formation. Da dieser aber wie der restliche Cortex einen 6-schichtigen Aufbau hat, gehört er eigentlich zum Cortex des Temporallappens. Der entorhinale Cortex ist die Eingangspforte des Hippocampus (und damit natürlich funktionell eng mit ihm verbunden). Hier laufen nahezu alle Informationen ein, die die hippocampale Formation aus dem Thalamus, dem Gyrus cinguli, der Amygdala und nahezu allen Cortexarealen erhält. Das Septum und einige Neurotransmitter zeigen aber auch eine direkte Innervation der einzelnen hippocampalen Strukturen. Vom entorhinalen Cortex wird die Information sowohl über den so genannten Tractus perforans an den Gyrus dentatus also auch direkt an den Hippocampus proper weitergegeben. Auch der Gyrus dentatus projiziert über die Moosfasern zum Hippocampus proper. Die Ausgänge aus dem Hippocampus laufen dann nahezu alle über das Subiculum. Von dort zieht der Fornix als das Hauptfaserbündel des Hippocampus zu den Mamillarkörperchen im Hypothalamus, um in den Papez-Kreislauf einzugehen (s.o.). Vom Fornix gehen aber auch direkte Abzweigungen des Subiculums zum Septum und zur Amygdala ab.
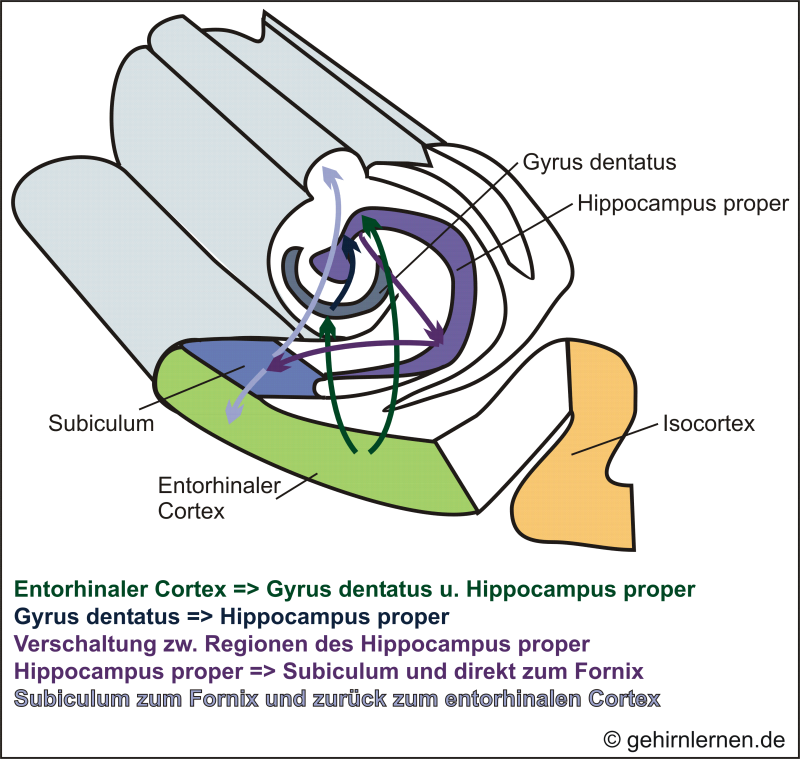 Abb. 5.3: Schaltkreis im Hippocampus (Querschnitt, schematisch)
Abb. 5.3: Schaltkreis im Hippocampus (Querschnitt, schematisch)
Schneidet man quer durch den Hippocampus hindurch, kann man die unterschiedlichen Strukturen sichtbar machen und die Verbindungen zwischen ihnen verdeutlichen: Der entorhinale Cortex projiziert über den Tractus perforans zum Gyrus dentatus aber auch direkt zum Hippocampus proper. Die Axone der Gyrus-dentatus-Zellen ziehen ebenfalls zum Hippocampus proper. Der Hippocampus projiziert sowohl über das Subiculum als auch direkt in den Fornix und über diesen zu den Mamillarkörperchen des Hypothalamus und damit in den Papez-Kreislauf.
Funktion
Durch seine Eingänge aus allen Assoziationsgebieten des Cortex weiß der Hippocampus – salopp gesagt – immer, was im Bewusstsein gerade vor sich geht. Dazu gehört es insbesondere zu wissen, wo man gerade ist. Im Hippocampus liegt das Ortsgedächtnis. Dafür gibt es im Hippocampus proper sogenannte Ortszellen, d.h. Pyramidenzellen, die für einen bestimmten Ort stehen. Man hat festgestellt, dass Tiere, die ihr Fressen verstecken (wie z.B. das Eichhörnchen), größere Hippocampi haben als Tiere, die von der Hand in den Mund leben. Auch bei z.B. Taxifahrern, die sich besonders gut orientieren können müssen, sind die Hippocampi größer als bei anderen Menschen.
Darüber hinaus ist der Hippocampus für die Gedächtnisbildung von Bedeutung. Im Hippocampus fließen Eingänge verschiedenster sensorischer Gebiete zusammen, werden verarbeitet und dann in den Cortex zurückgesandt (siehe oben Papez-Kreislauf), wo sie dauerhaft gespeichert werden können. Durch den Hippocampus findet dabei eine Überführung von Gedächtnisinhalten aus dem Kurz- in das Langzeitgedächtnis statt. Diese nennt man Gedächtniskonsolidierung. Sind bei einem Menschen beide Hippocampi zerstört oder entfernt (z.B. bei schwerster Epilepsie), so ist sein Langzeitgedächtnis zwar intakt (weil der Cortex intakt ist), neue Inhalte können aber nicht mehr abgespeichert werden.
Dabei agiert der Hippocampus auch wie ein Neuigkeitsdetektor. Er registriert, wenn eine Information neu präsentiert wurde, um diese dann für die Speicherung vorzubereiten. Ist die Information bereits bekannt, so kann er sich diese Arbeit „sparen“. Wird die gleiche Information leicht variiert oder in einem anderen Kontext präsentiert, so kann der Hippocampus die vorhandene Gedächtnisspur abrufen und reaktivieren, um sie dann mit der neuen Verknüpfung erneut abzulegen. So werden mit jedem Erinnern abgelegte Gedächtnisinhalte reaktiviert und weiterverarbeitet. Dies führt zum einen zu einer durch die Verknüpfung stabileren Gedächtnisspur, zum anderen aber auch bei jedem Abruf zu einer leicht veränderten Erinnerung (So kann man sich erklären, dass sich Geschichten, die von derselben Person immer wieder erzählt werden, immer leicht verändern und weiterentwickeln).
Durch seine starken Verbindungen zur Amygdala, dem Furchtzentrum unseres Gehirns, kommt dem Hippocampus eine zusätzliche Rolle bei der Verarbeitung von Emotionen zu. Zusammen mit der Amygdala formt er ein emotionales Gedächtnis. Dies macht den Hippocampus aber auch anfällig für starke emotionale Reize und Stressoren. So führen Traumata zu einer Verkleinerung des Hippocampus (Atrophie) ebenso wie Depressionen, Angst- und Essstörungen.
Plastizität: Langzeitpotenzierung, Synaptogenese und Neurogenese
Um all diesen – stetig wechselnden – Aufgaben gerecht werden zu können, besitzt der Hippocampus ein hohes Maß an Plastizität, d.h. an Fähigkeit sich selbst zu verändern und anzupassen. Dazu gehört die Möglichkeit, dass sich die Stärke der synaptischen Übertragung zwischen zwei Zellen verringern oder verstärken kann. Dabei verstärken sich viel genutzte Verbindungen (Langzeitpotenzierung, LTP) und wenig genutzte schwächen sich ab (Langzeitdepression, LTD). Darüber hinaus wurde gezeigt, dass sich nach dem gleichen Prinzip neue Verbindungen zwischen Nervenzellen ausbilden und alte nicht mehr benutzte Verbindungen abgebaut werden können. Nirgendwo anders im Gehirn ist diese Fähigkeit zur Synaptischen Plastizität oder Synaptogenese so stark ausgeprägt wie im Hippocampus. Angetrieben wird diese Synaptogenese zudem durch die Fähigkeit zur Bildung neuer Nervenzellen. Im Säugergehirn gibt es nur zwei Regionen, die diese Fähigkeit überhaupt noch besitzen, eine liegt im Riechhirn und eine im Hippocampus, genauer gesagt im Gyrus dentatus. Diese neue Nervenzellen machen den Hippocampus so flexibel in der Aufnahme und Verarbeitung von neuen Reizen und treiben zudem, da neue Zellen natürlich auch Kontakte im vorhandenen Zellnetzwerk ausbilden wollen, die Bildung neuer Synapsen an. Insbesondere durch diese hohe Flexibilität ist der Hippocampus aber auch anfällig für schädliche äußere Reize (s.o.).
Der Hypothalamus
Der Hypothalamus liegt im Zwischenhirn direkt unterhalb des Thalamus und in unmittelbarer Nachbarschaft zum Seitenventrikel (vgl. Abb. 2.2). Seinen Namen trägt er von der Lage her: das griechische „hypo“ bedeutet zu Deutsch „unter“.
Der Hypothalamus ist die Steuerzentrale des inneren Milieus, d.h. der Homöostase. Er reguliert die Schilddrüsenfunktion, die Körpertemperatur, das Wachstum, den Schlaf-Wach-Rhythmus, die innere Uhr, den Appetit, die Sättigung, den Energiehaushalt, das Körpergewicht, den Salz- und Wasserhaushalt und den Sexualtrieb. Dabei können Zellen des Hypothalamus den Zustand von Blut und Liquor messen und über Verbindungen zum vegetativen Nervensystem und die Hormonausschüttung regulierend eingreifen. Letzteres geschieht insbesondere durch das Zusammenspiel mit der Hirnanhangdrüse, der Hypophyse, die dem Hypothalamus – salopp gesagt – anhängt. Auf „Befehl“ des Hypothalamus, der in Form von Releasing-Hormen die Hypophyse erreicht, schüttet diese stimulierende Hormone ins Blut aus, die über den Blutkreislauf in die Drüsen gelangen, in denen daraufhin die eigentlichen "Ziel"-Hormone ausgeschüttet werden.
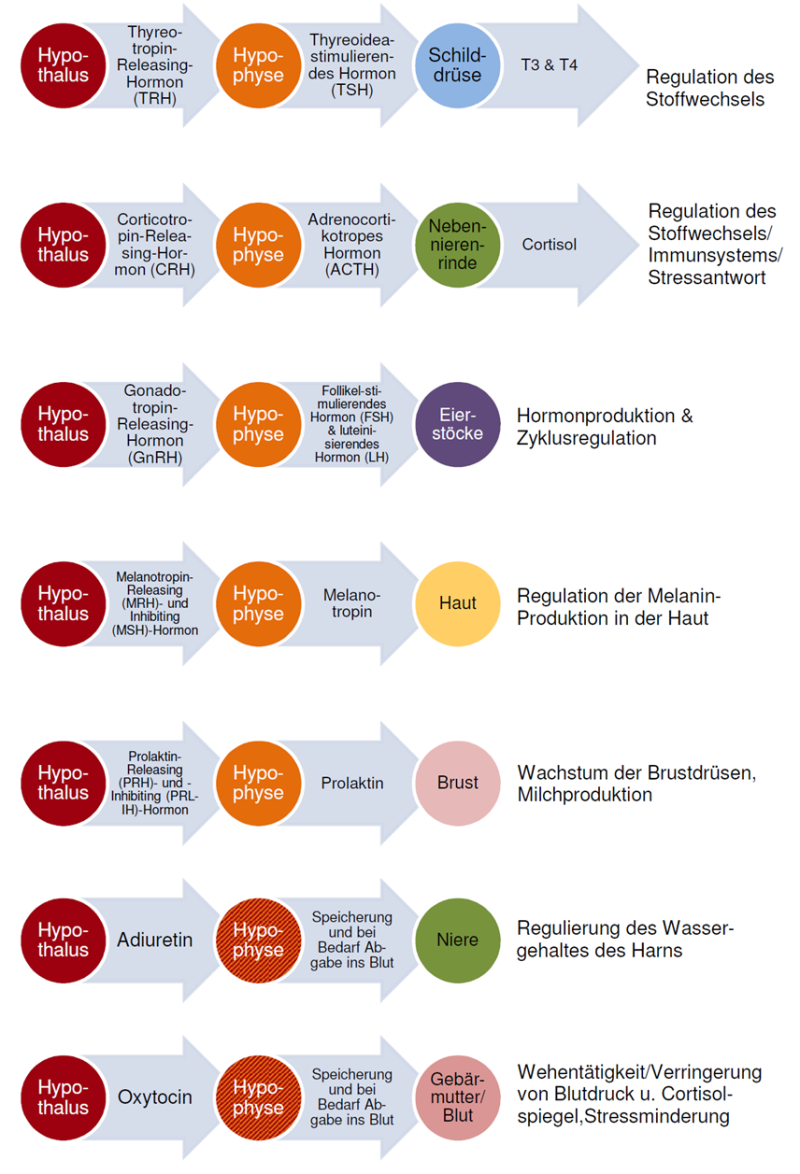 Abb. 5.4: Regulation einiger wichtiger Hormone über Hyothalamus und Hypophyse (© gehirnlernen.de)
Abb. 5.4: Regulation einiger wichtiger Hormone über Hyothalamus und Hypophyse (© gehirnlernen.de)
Darüber hinaus ist der Hypothalamus über die Formatio reticularis an der übergeordneten Steuerung von z.B. Herz-Kreislauffunktionen beteiligt. Durch seine Funktionen der hormonellen Steuerung, seine engen Verbindungen mit dem limbischen System und Einbezogenheit in den Papez-Kreislauf (siehe oben) wird der Hypothalamus auch mit zum limbischen System gezählt. Die Amygdala (siehe unten) nimmt mit direkten Verbindungen zum Hypothalamus Einfluss auf die Ausschüttung von Hormonen im Zuge der Furchtreaktion.
Der Gyrus cinguli
Der Gyrus cinguli ist ein innenliegender Teil des Cortex. Er liegt wie ein Gürtel auf dem Balken auf und verläuft mit ihm von vorn nach hinten. Daher trägt der Gyrus cinguli, der auch kurz Cingulum genannt wird, seinen Namen, der auf Deutsch Gürtelwindung heißt. Betrachtet man den Gyrus cinguli auf Zellebene, so fällt auf, dass sich zwei Bereiche unterscheiden lassen: der vordere Teil (pars anterior) und der hintere Teil (pars posterior). Nach den Funktionen können vom vorderen Teil auch noch die Area subcallosa (unter dem Balken liegend) und die cingulären Motorareale abgrenzen.
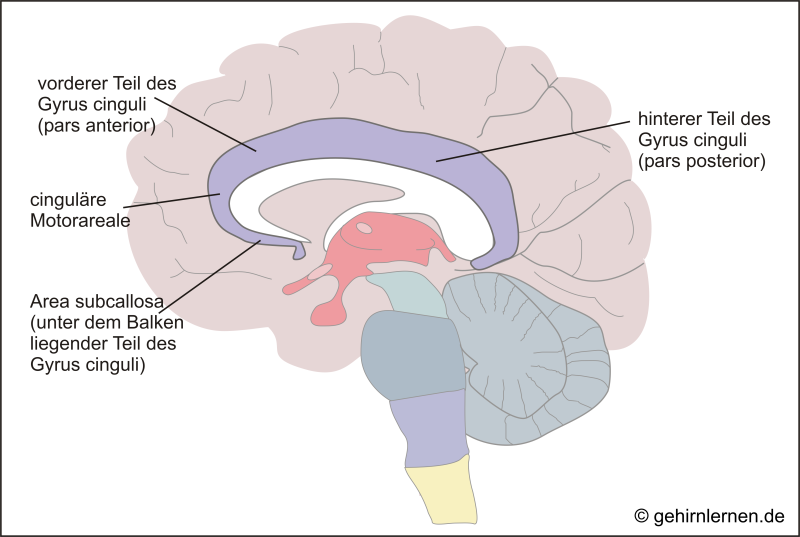 Abb. 5.5: Gyrus cinguli
Abb. 5.5: Gyrus cinguli
Der hintere Teil des Cingulums ist mit dem Parietal-, Temporal-, und Frontallappen eng verbunden. Durch die Regulation von visuell-räumlicher Aufmerksamkeit ist er am räumlichen Gedächtnis (im Zusammenspiel mit dem Hippocampus) beteiligt.
Der vordere Teil ist eng mit der Amygdala, dem Hippocampus, dem Nucleus accumbens, dem Thalamus und dem insulären Cortex verbunden. Seine Aufgabe wird vor allem darin gesehen, widersprüchliche Reize gegeneinander abzuwägen und eine Entscheidung zu treffen.
 Abb. 5.6: Der Stroop-Test (Stroop, 1935)
Abb. 5.6: Der Stroop-Test (Stroop, 1935)
Beim Stroop-Test werden dem Probanden Farbwörter in widersprüchlichen Farben gezeigt. Die Schwierigkeit liegt dabei, dass die linke Gehirnhälfte das Wort lesen, die rechte die Farbe benennen will. Diese widersprüchlichen Informationen werden im Gyrus cinguli verarbeitet und es wird eine Entscheidung für das eine oder das andere getroffen.
Über seine Verbindung mit dem insulären Cortex ist er zudem an der Schmerzbewertung beteiligt.
Über die cingulären Motorareale ist der Gyrus cinguli schließlich in motorische Funktionen – insbesondere Mimik und Gestik – eingebunden.
Das Cingulum ist eine Schaltstelle des Papez-Kreislaufes und damit Teil des limbischen Systems (siehe oben).
Die Amygdala - der Mandelkern
Die Amygdala ist eine mandelförmige Struktur ganz vorn im Schläfenlappen vor dem Hippocampus (siehe Abb. 5.1). Ihren Namen trägt sie von ihrer Form her: Amygdala wurde aus dem griechischen Wort für Mandel abgeleitet. Zu Deutsch heißt sie darum Mandelkern.
Die Amygdala spielt eine wichtige Rolle in der Bildung von Emotionen, insbesondere von Furcht und Angst. Zudem ist sie an der Abspeicherung von emotional besetzten Erinnerungen beteiligt.
Genau genommen ist die Amygdala ähnlich wie der Hippocampus keine einheitliche Struktur, sondern ein Kernkomplex aus ca. 13 Kernen, die ihren entwicklungsgeschichtlichen Ursprung teils im Cortex, teils im Striatum haben. Die Verbindungen und Funktionen dieser einzelnen Kerne sind noch nicht alle erforscht. Gemeinhin hat man sich auf drei bis vier wichtige Kerngruppen geeinigt:
- Zentraler Komplex
- Basolateraler Komplex, von dem manche den lateralen Komplex gesondert betrachten (so auch in Abb. 5.6)
- Corticale Kerngruppe
Betrachtet man die Abbildung 5.6, so scheinen die Verbindungen zur, von und innerhalb der Amygdala zunächst sehr komplex. Es lassen sich aber Haupt- (dicke Pfeile) und Nebenverbindungen (dünnere Pfeile) unterscheiden.
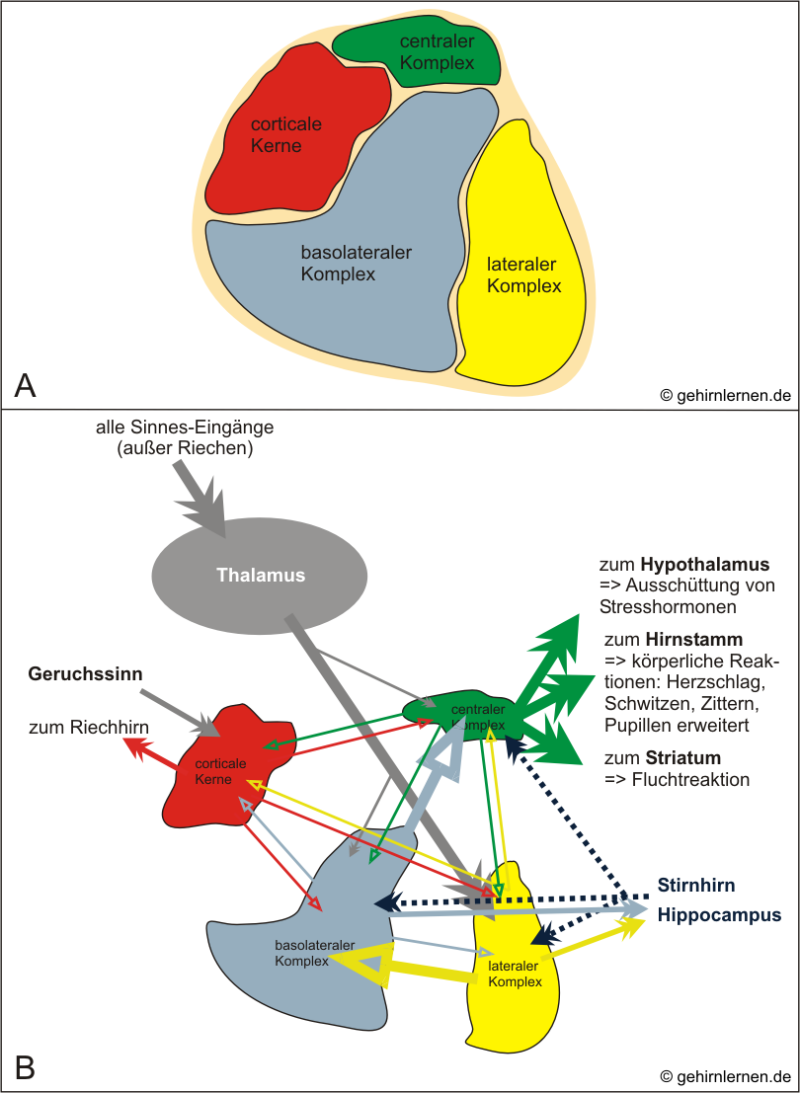 Abb. 5.7: Die Amygdala und ihre Verschaltungen
Abb. 5.7: Die Amygdala und ihre Verschaltungen
A) Anordnung der Kernkomplexe in der menschlichen Amygdala im Querschnitt (Frontalschnitt). B) Informationsfluss und interne Verschaltungen in der Amygdala (Kernkomplexe zur Veranschaulichung auseinandergezogen - für ausführliche Erläuterung siehe Text).
Der laterale Komplex ist die Haupteingangspforte der Amygdala. Die meisten Sinneseingänge, die vom Thalamus zur Amygdala weitergeleitet werden, kommen in diesem Komplex an. Zur weiteren Verarbeitung gibt der laterale Komplex die Information an den basolateralen Komplex weiter, der wiederum hauptsächlich zum zentralen Komplex projiziert. Der zentrale Komplex stellt die Ausgangspforte der Amygdala dar. Von ihm aus werden über Projektionen zum Hirnstamm körperliche Reaktionen wie Zittern, Anspannung, erweiterte Pupillen, Schwitzen, über Projektionen zum Hypothalamus die hormonelle Stressantwort und über Projektionen zum Striatum die Fluchtreaktion generiert.
Dieser Hauptweg der Verarbeitung in der Amygdala gilt für alle Sinneswahrnehmungen wie Sehen, Hören, Fühlen, nicht aber für das Riechen. Über ihre corticalen Kerngebiete erhält die Amygdala Geruchsinformationen direkt und ohne Vorarbeitung durch den Thalamus. Die corticalen Kerngebiete projizieren auch zurück zum Riechhirn.
Die Amygdala ist zwar das entscheidende Zentrum für die Furchtverarbeitung (fehlt sie, hat der Betreffende auch keine Furcht oder Angst mehr), doch arbeitet auch sie nicht lösgelöst von den anderen Gehirnbereichen. Alle Komplexe der Amygdala – abgesehen von den corticalen Kernen – erhalten modulierende Eingänge aus dem Stirnhirn und dem Hippocampus.
Dabei übt das Stirnhirn eine übergeordnete Kontrollfunktion über die Amygdala aus. So können Emotionen und Angst zu einem gewissen Grade unterdrückt und kontrolliert werden. Die Verbindung des Stirnhirns zur Amygdala ist aber nicht so stark wie die der Amygdala zum Stirnhirn, so dass Angst nicht gänzlich unterdrückt werden kann.
Durch das Zusammenspiel von Amygdala und Hippocampus wird ein emotionales Gedächtnis aufgebaut. Einen schweren Unfall z.B. wird man sein Leben lang nicht mehr vergessen und, kehrt man an den Ort des Unfalls zurück, so werden durch das Zusammenspiel von Hippocampus und Amygdala die Erinnerungen wieder wach und möglicherweise sogar erneut körperliche Reaktionen ausgelöst.
Angst ist eine lebenswichtige Funktion. Zwar werden Angst und Furcht in unserer heutigen Gesellschaft häufig als Makel oder Schwäche angesehen, eigentlich sind sie aber lebenswichtige Reaktionen, die uns vor Gefahr schützen sollen. Darum ist es auch sinnvoll, dass sie nie ganz vom Stirnhirn unterdrückt werden können. Ein weiteres Indiz für die Wichtigkeit der Angst ist auch, dass eingehende Sinnesreize nicht erst kompliziert über den Thalamus an die zuständigen somatosensorischen Rindenbereiche weitergegeben, dort verarbeitet und dann an die Amygdala weitergeleitet werden, sondern auch direkt vom Thalamus ohne Vorverarbeitung – d.h. auch nur als vages Abbild – an die Amygdala geschickt werden.
Stellen wir uns z.B. einmal vor, wir durchwandern wie unsere menschlichen Vorfahren die afrikanische Steppe. Plötzlich fällt der Anblick einer Schlange in unser Auge. Das Bild wird an den Thalamus weitergegeben. Dieser schickt es einerseits natürlich zur Sehrinde, aber andererseits auch – wenn auch nur als ungenaues, unbearbeitetes Bild – sofort zur Amygdala. Diese löst eine Furchtreaktion aus und lässt uns zurückspringen. Jetzt kommt das Bild auch über die Sehrinde im Bewusstsein an. Die Sehrinde schickt das überarbeitete Bild zum Abgleich zur Amygdala: ist es doch nur ein Seil, so wird die Reaktion der Amygdala wieder heruntergefahren; ist es aber wirklich eine Schlange, so wäre diese Information von der Sehrinde möglicherweise schon zu spät gekommen: die Schlange hätte uns schon gebissen – gut, dass die Amygdala schon früher reagieren konnte!
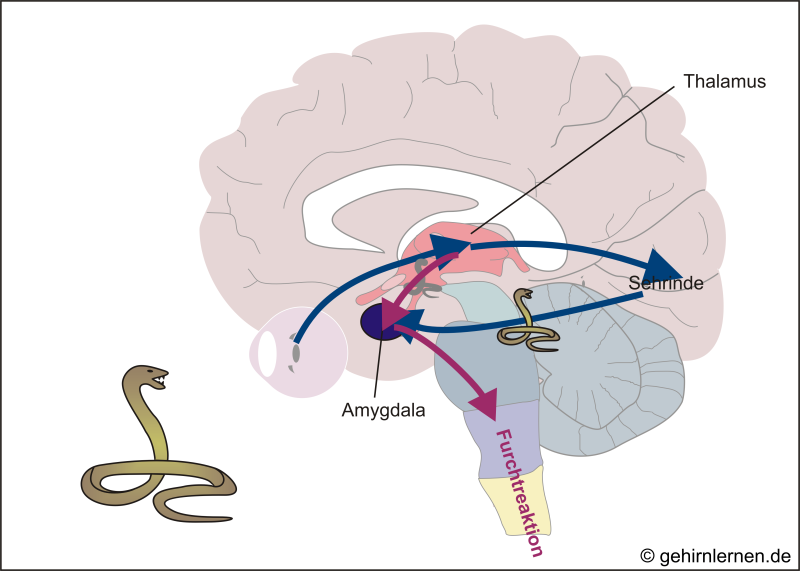 Abb. 5.8: Furchtreaktion - Was im Gehirn passiert (Erläuterung im Text)
Abb. 5.8: Furchtreaktion - Was im Gehirn passiert (Erläuterung im Text)
Zusammengefasst: die Furchtreaktion ist schneller als die bewusste Wahrnehmung des Auslösers – ein oftmals lebenswichtiger Zeitgewinn!
Der Nucleus accumbens
An kaum einem anderen Kerngebiet zeigt sich die Schwierigkeit der Begrenzung des limbischen Systems besser als am Nucleus accumbens. Aufgrund seiner Zugehörigkeit zum „Belohnungssystem“ ist es gerechtfertigt ihn funktionell zum limbischen System zu rechnen. Anatomisch gesehen ist er aber ein Teil des Striatums, das er bauchwärts begrenzt. Dies trug ihm auch den Namen „ventrales Striatum“ ein.
Der Nucleus accumbens besteht aus einer Schalenregion (shell) und einer Kernregion (core). Die Kernregion stellt eine Art ventrale Erweiterung des Striatums dar und ist vor allem mit dem motorischen System verbunden, zu dem es projiziert. Die Schalenregion hingegen unterhält hauptsächlich Kontakte zum limbischen System (v.a. Hippocampus, Amygdala), wobei es von dort Projektionen erhält.
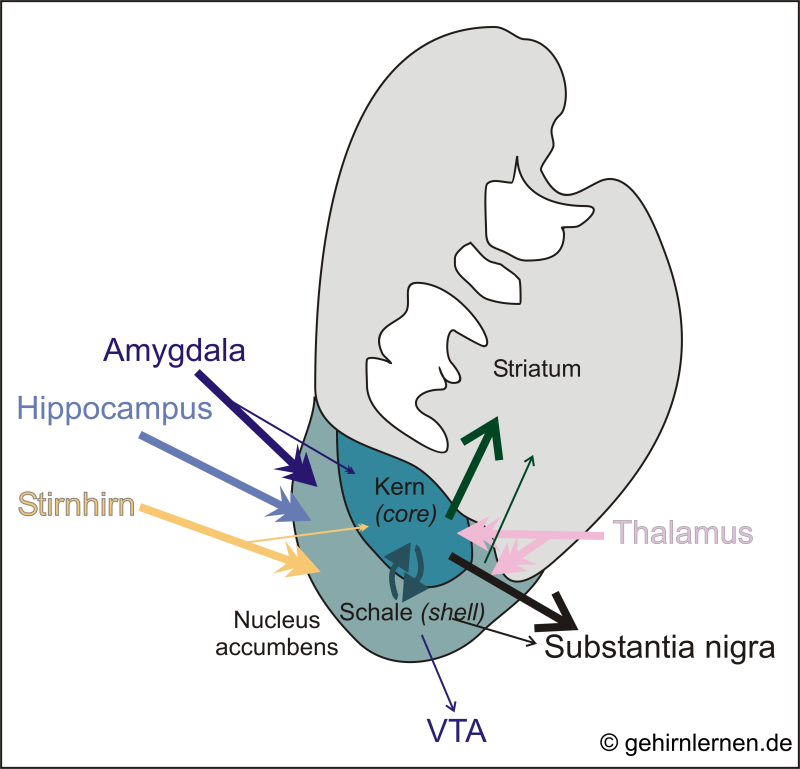 Abb. 5.9: Verschaltungen des Nucleus accumbens
Abb. 5.9: Verschaltungen des Nucleus accumbens
Die meisten Eingänge erhält der Nucleus accumbens über die Schalenregion aus limbischen Arealen und dem Stirnhirn. Diese Regionen projizieren auch geringfügig in die Kernregion. Der Thalamus projiziert gleichermaßen zu beiden Regionen. Die hauptsächlichen Ausgänge des Nucleus accumbens laufen über die Kernregion, die eng mit dem Striatum verbunden ist, zu dem sie auch vornehmlich projiziert. Über die Substantia nigra und die VTA wird überdies die Dopaminausschüttung in das Striatum moduliert (dicke Pfeile markieren starke, dünne Pfeile schwache Projektionen).
Diese limbischen Eingänge und motorischen Ausgänge machen den Nucleus accumbens zu einer funktionellen Schnittstelle zwischen dem limbischen und dem motorischen System (limbic-motoric interface). Dabei ist es seine Aufgabe motivationales Verhalten in Handlung umzusetzen. Darüber hinaus ist er an emotionalen Lernprozessen, Empathie und Freundschaft, Sympathie und Präferenz beteiligt.
Stimulationsexperimente mit dem Nucleus accumbens brachten ihm den Namen „Belohnungssystem“ ein. Die Stimulation des Nucleus accumbens erzeugt nämlich Glücksgefühle. Ratten, denen James Olds und Peter Milner 1954 Elektroden in den Nucleus accumbens implantierten, damit sie sich selbst dort elektrisch stimulieren konnten, zeigten sich so verzückt, dass sie alles andere – von Essen bis Sexualkontakten – außer Acht ließen und in kürzester Zeit von der Selbststimulation abhängig wurden. Aufgrund der hohen Innervationsdichte von Dopamin aus der VTA im Nucleus accumbens haben Drogen, die auf Dopamin verstärkend wirken (Amphetamine, Kokain, Cannabis, Ketamin („special K“) ebenso euphorisierende aber auch ebenso schnell süchtig machende Wirkung. Suchtverhalten wird somit auch über den Nucleus accumbens realisiert.